Rôle du muscle squelettique strié dans la conversion de l’énergie
I. Enregistrement de la contraction musculaire
1. Notion de potentiel de repos et potentiel d'action
Le potentiel de repos ou potentiel membranaire est la différence de tension mesurée entre la face externe et la face interne de la cellule.
C'est la différence de concentration ionique de part et d'autre de la membrane qui détermine la valeur du potentiel transmembranaire. Il est de $-70 \text{ mV}$.
La stimulation électrique de la cellule provoque une brusque modification du potentiel de repos. C'est un phénomène électrique qui naît suite à l'excitation et qu'on appelle potentiel d'action. Il est composé de :
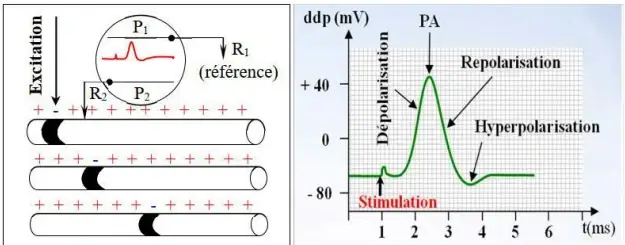
- L'artefact de stimulation : phénomène électrique enregistré par les électrodes réceptrices, indiquant le moment de stimulation.
- La dépolarisation : le potentiel augmente très rapidement (phase ascendante) pour atteindre un pic.
- La repolarisation : Après le pic, le potentiel diminue pour se rapprocher du potentiel de repos.
- L'hyperpolarisation : Le potentiel devient inférieur au potentiel de repos.
2. Réponse du muscle aux excitations électriques
a) Cas d'une excitation unique
Lorsqu'on applique sur le muscle une excitation électrique unique et efficace, on obtient une contraction brève et isolée à laquelle on donne le nom de secousse musculaire.
Le myogramme (M) obtenu est composé de trois phases :
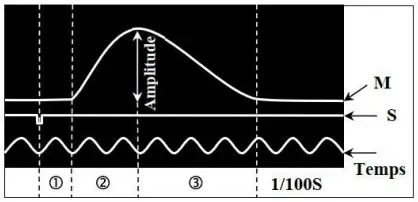
- La phase de latence ① : durée entre le moment de l'excitation et le début de la réponse du muscle.
- La phase de contraction ② : au cours de laquelle la longueur du muscle décroit (muscle se raccourcit).
- La phase de relâchement ③ : au cours de laquelle le muscle reprend ses dimensions initiales (sa durée est légèrement supérieure à celle de la phase de contraction).
b) Cas de plusieurs excitations à intensité croissante
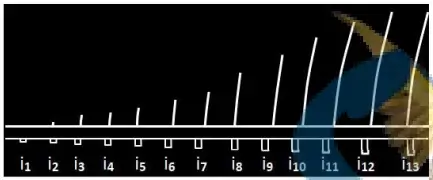
- L'excitation ($i_1$) : pas de réponse, $i_1$ est donc inefficace (excitation infraliminaire).
- À partir de ($i_2$) (Seuil d'excitation = rhéobase), on enregistre une réponse dont l'amplitude augmente avec l'augmentation de l'intensité, c'est la loi de recrutement (augmentation du nombre d'unités musculaires recrutées).
- À partir de ($i_{12}$), l'amplitude de la réponse reste constante même si l'intensité de l'excitation continue d'augmenter, car toutes les unités constituant le muscle se contractent.
c) Cas de deux excitations rapprochées
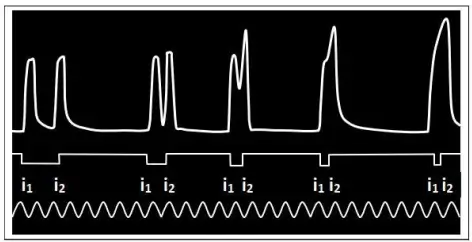
On soumet plusieurs fois un muscle à deux excitations efficaces successives de même intensité ($i_1, i_2$). À chaque fois, on diminue l'intervalle de temps entre les deux excitations. La réponse diffère selon l'instant où on applique la deuxième excitation :
- Si les deux excitations sont suffisamment éloignées, on enregistre deux secousses musculaires isolées et de même amplitude.
- Si les deux excitations sont rapprochées et que la 2ème excitation atteint le muscle pendant la phase de relâchement de la réponse précédente, il se produit une fusion incomplète (partielle) des deux secousses musculaires avec une augmentation de l'amplitude de la 2ème secousse.
- Si les deux excitations sont très rapprochées et que la 2ème excitation atteint le muscle pendant la phase de contraction de la réponse précédente, on observe une fusion complète (totale) des deux secousses qui apparaissent comme s'il n'y avait qu'une seule secousse musculaire d'une amplitude plus grande.
d) Cas d'une série d'excitations successives
On soumet le muscle à une série d'excitations efficaces de même intensité et en variant la fréquence des excitations :
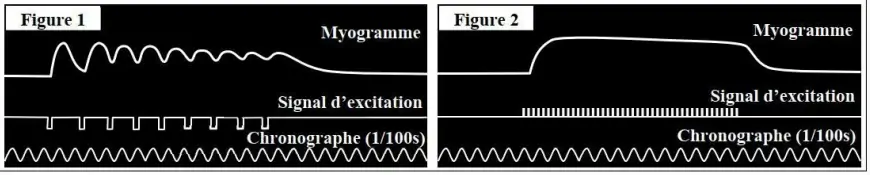
Figure 1 : quand la fréquence des excitations est faible, le myogramme obtenu prend l'allure d'un palier sinueux. La réponse du muscle est dite, alors, tétanos imparfait. Ce phénomène peut être expliqué par la fusion incomplète des secousses musculaires, car chaque excitation atteint le muscle pendant la phase de relâchement de la réponse précédente.
Figure 2 : quand la fréquence des excitations est forte, le myogramme obtenu prend l'allure d'un palier droit. La réponse du muscle est dite, alors, tétanos parfait. Ce phénomène peut être expliqué par la fusion complète des secousses musculaires, car chaque excitation atteint le muscle pendant la phase de contraction de la réponse précédente.
e) Effet de la fatigue sur la contraction musculaire
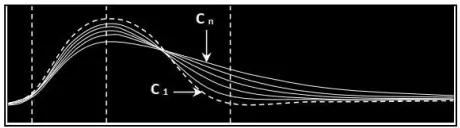
On applique une série d'excitations de même intensité pendant une durée très longue. $C_1$ = la secousse obtenue à la suite de la première excitation et $C_n$ = la secousse obtenue à la suite de la dernière excitation.
La fatigue musculaire se manifeste donc par la diminution de l'amplitude de la réponse musculaire et par une augmentation du temps de relâchement.
II. Les phénomènes accompagnant la contraction musculaire
1. Les phénomènes thermiques
Pour mesurer la chaleur dégagée lors de la contraction musculaire, on utilise une thermopile.
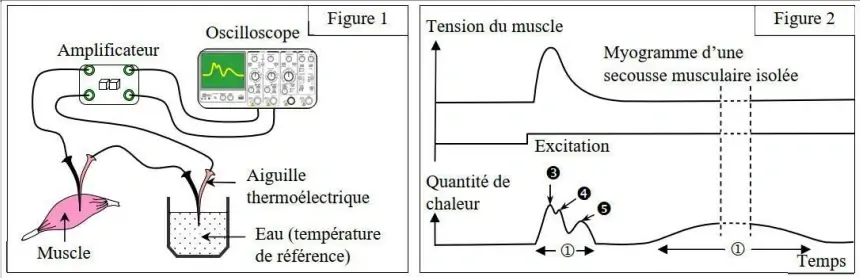
Au cours d'une activité musculaire, le muscle dégage de la chaleur en deux temps :
- La chaleur initiale (①), qui se dégage rapidement au cours de la secousse musculaire et dont une partie est libérée au cours de la phase de contraction (Chaleur de contraction (③) et chaleur de soutient (④)), et l'autre partie au cours de la phase de relâchement (Chaleur de relâchement (⑤)).
- La chaleur retardée (②) se dégage lentement après la secousse musculaire.
2. Les phénomènes chimiques et énergétiques
Lors de l'activité musculaire, le muscle :
- Bénéficie d'une augmentation du débit sanguin qui permet l'intensification des échanges ;
- Utilise beaucoup plus de glucose et du dioxygène, et produit davantage de $CO_2$ ;
- Ne consomme pas les protides et les lipides mais utilise uniquement le glucose que ce soit en activité ou au repos.
Ces phénomènes traduisent l'oxydation du glucose qui produit l'énergie nécessaire à la contraction musculaire.
III. Structure et ultrastructure du muscle squelettique strié
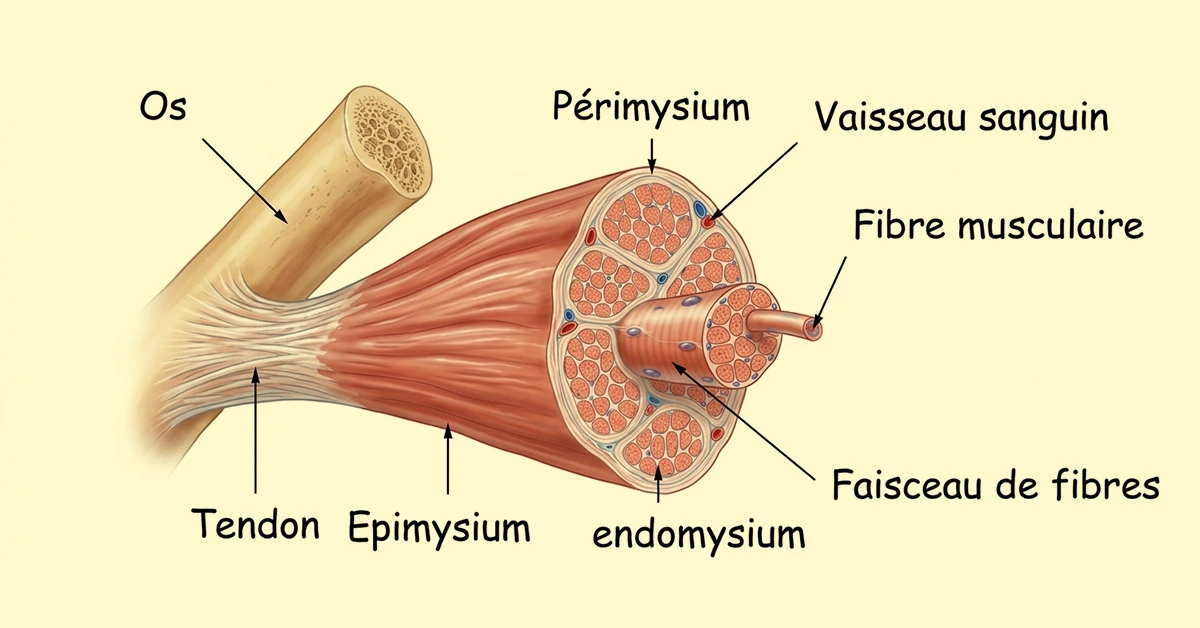
1. Structure du muscle squelettique strié
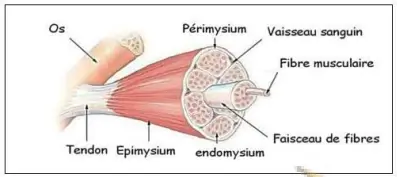
Le muscle squelettique est enveloppé par diverses membranes. Une qui entoure le muscle c'est l'épimysium. Ensuite, le muscle est divisé en faisceaux de fibres qui sont séparés les uns des autres par le périmysium. Enfin les faisceaux sont composés de fibres musculaires entourées par l'endomysium. Aux extrémités du muscle, les différentes membranes conjonctives s'unissent pour former les tendons qui attachent les muscles aux os.
2. L'ultrastructure de la fibre musculaire
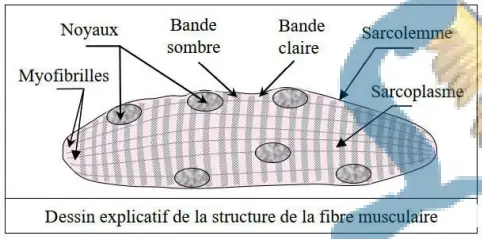
La fibre musculaire est une cellule qui possède plusieurs noyaux (cellule plurinucléée = syncytium), limités par une membrane (sarcolemme). Chaque fibre présente une striation transversale (alternance de bandes sombres et de bandes claires), d'où le nom de muscle squelettique strié.
Le cytoplasme de la fibre musculaire (ou sarcoplasme) contient des structures longitudinales parallèles : les myofibrilles.
- Chaque myofibrille est constituée de myofilaments. On distingue deux types de myofilaments :
- Des myofilaments fins d'actine, qui sont disposés de part et d'autre des stries Z.
- Des myofilaments épais de myosine, se situent entre les myofilaments fins.
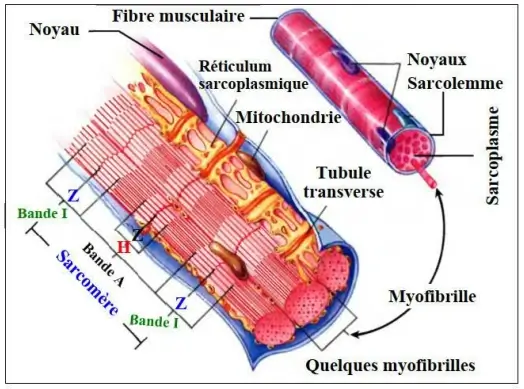
- L'espace entre deux stries Z est le sarcomère.
- L'endroit où il n'y a que les myofilaments épais de myosine est appelé zone H.
- La zone où il n'y a que les myofilaments fins est appelée la bande I.
- La zone où il y a les myofilaments fins et les myofilaments épais est appelée la bande A.
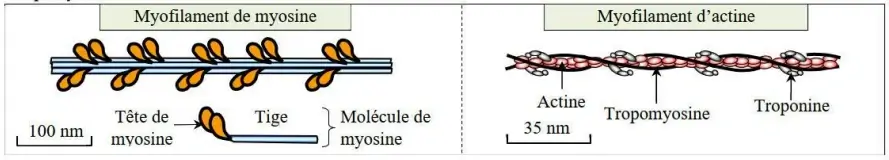
- Le myofilament épais est composé de protéine myosine (2 têtes), alors que le myofilament fin est composé des protéines actine + troponine + tropomyosine.
IV. Mécanisme de la contraction musculaire
1. En quoi consiste la contraction musculaire ?
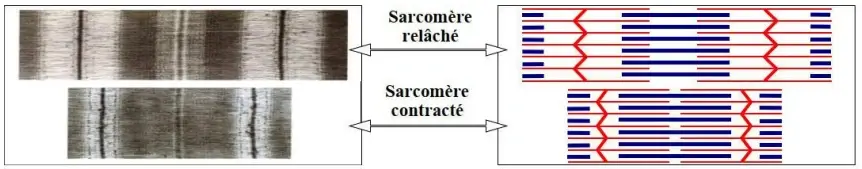
Lors de la contraction musculaire on observe :
- Un raccourcissement des sarcomères (rapprochement des stries Z).
- Une réduction de la longueur des bandes claires et de la bande H.
- Une constance des bandes sombres.
- La longueur des myofilaments reste constante.
Ceci prouve qu'il y a, au cours de la contraction, un glissement des myofilaments d'actine par rapport aux myofilaments de myosine. Le sarcomère est donc l'unité fonctionnelle de la fibre musculaire.
2. Mécanisme du glissement des myofilaments
a) Rôle du calcium dans la contraction musculaire
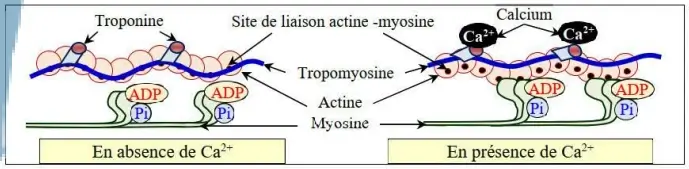
En absence des ions $Ca^{2+}$ (au repos), la tropomyosine cache le site de fixation de la tête de myosine sur l'actine. La fixation des ions $Ca^{2+}$ sur la troponine entraîne le déplacement de la tropomyosine ce qui permet de démasquer le site de fixation de la tête de myosine sur l'actine, et par suite, la fixation de myosine sur l'actine et la formation des complexes actomyosines.
b) Mécanisme du glissement des myofilaments
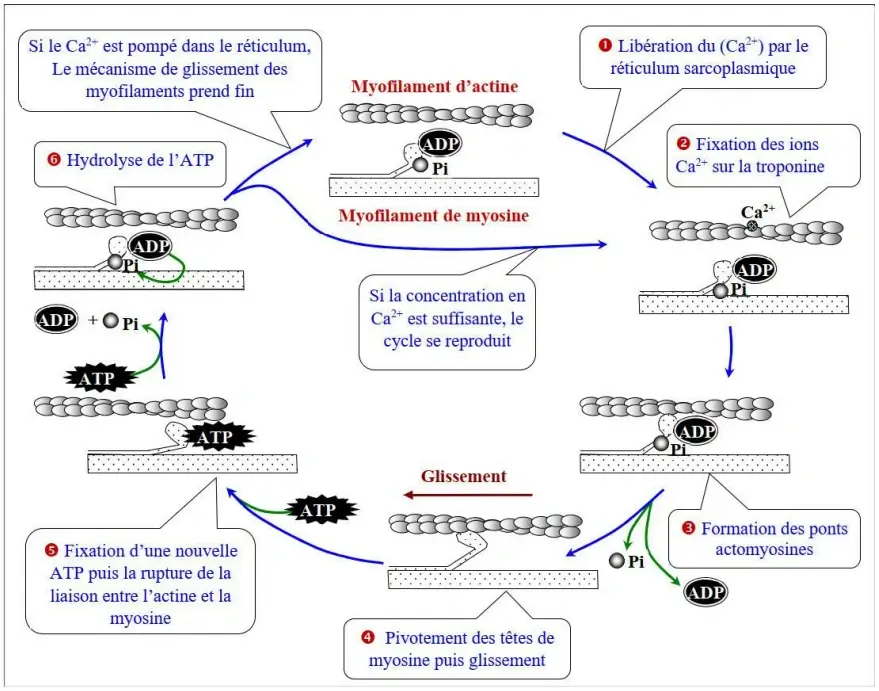
- Libération du $Ca^{2+}$ par le réticulum sarcoplasmique.
- Fixation des ions $Ca^{2+}$ sur la troponine.
- Formation des ponts actomyosines.
- Pivotement des têtes de myosine puis glissement.
- Fixation d'une nouvelle molécule d'ATP puis la rupture de la liaison entre l'actine et la myosine.
- Hydrolyse de l'ATP : $ATP \rightarrow ADP + P_i$.
Si le $Ca^{2+}$ est pompé dans le réticulum, le mécanisme de glissement des myofilaments prend fin. Si la concentration en $Ca^{2+}$ est suffisante, le cycle se reproduit.
V. Les voies de régénération de l'ATP au cours de la contraction musculaire
a) Voies anaérobies immédiates
- La voie de la phosphocréatine : La production d'ATP par hydrolyse de la phosphocréatine. $$PC + ADP \xrightarrow{\text{Créatine kinase}} C + ATP$$
- La voie de l'ADP : La production d'ATP par phosphorylation d'ADP. $$2 ADP \xrightarrow{\text{Myokinase}} ATP + AMP$$
b) Voies anaérobies de moyenne vitesse : La fermentation lactique
$$C_6H_{12}O_6 \longrightarrow 2 CH_3\text{-}CHOH\text{-}COOH + 2 ATP$$Glucose → Acide lactique
c) Voies aérobies lentes : l'oxydation respiratoire
$$C_6H_{12}O_6 + 6 O_2 + 38 ADP + 38 P_i \longrightarrow 6 CO_2 + 6 H_2O + 38 ATP$$Glucose → Énergie + Déchets
Les différents types de fibres musculaires
Caractéristiques des différents types de cellules musculaires
| Propriétés | Muscle squelettique | Muscle cardiaque | Muscle lisse |
|---|---|---|---|
| Caractéristiques cellulaires | Cellules très allongées et cylindriques avec de nombreux noyaux | Cellules irrégulières en forme de bâtonnet habituellement avec un seul noyau | Cellules fusiformes avec un seul noyau |
| Taille maximale des cellules (lg x d) | 30 cm × 100 μm | 100 μm × 15 mm | 500 μm × 5 mm |
| Noyaux | Multinucléée (périphériques) | Un seul au centre | Un seul au centre |
| Striations visibles | Oui | Oui | Non |
| Contraction | Volontaire | Involontaire | Involontaire |
| Innervation | Somatique | Autonome (sympathique et parasympathique) | Autonome (sympathique et parasympathique) |
| Localisation | Se fixe au squelette | Myocarde | Parois des viscères : vaisseaux sanguins |
Les fibres lentes ou fibres rouges ou de type I :
- Sont rouges car elles sont gorgées de sang. (Avoir un maximum d'$O_2$).
- Sont très fournies en mitochondries (Fabrication d'énergie par le biais d'$O_2$).
- Peu de nerfs les entourent car elles n'ont pas besoin de se contracter rapidement.
- Par contre, elles ont une forte capacité de résistance à l'effort.
- Sont fines et interviennent dans des exercices longs.
Les fibres rapides ou fibres blanches ou de type II :
- Sont pâles (Moins de sangs = moins d'$O_2$) ;
- Sont riches en réserves énergétiques (glycogène) et moins riches en mitochondries ;
- Ont un diamètre important ;
- Ont des capacités de contractions rapides et interviennent dans des mouvements brusques ;
- Leur innervation est importante pour que le signal arrive vite ;
- Ont une résistance faible à l'effort et ne sont pas capables de se contracter longtemps.
Effets de l'entraînement :
Généralement, chaque individu a une répartition égale de la part de chaque type de fibres mais certains ont des prédispositions.
L'entraînement peut modifier cette répartition de manière à avoir plus de fibres lentes ou plus de fibres rapides.
Ainsi, si vous vous entraînez pour un marathon, une partie de vos fibres rapides vont se transformer en fibres lentes pour permettre un effort long. À l'inverse, si vous faites un entraînement de sprinter ou d'haltérophile, une partie des fibres lentes va devenir rapide pour permettre d'aller plus vite et d'être plus fort. Quelqu'un qui ne fait pas de sport aura tendance à avoir plus de fibres rapides.